Р. Я. МЕШКОВАРУКОВОДСТВО ПО ИММУНОПРОФИЛАКТИКЕ ДЛЯ ВРАЧЕЙ
Иммунная система человека состоит из центральных (тимус, костный мозг) и периферических (селезенка, л/у) органов и работает как интегрированная защитная система организма, обеспечивая элиминацию инфекционных агентов и длительный протективный иммунитет. Микроорганизмы, окружающие человека ежедневно, у нормального здорового индивидуума только иногда могут быть причиной болезни. Большинство инфекционных агентов распознается и разрушается в течение нескольких часов благодаря защитным механизмам, которые не являются антигенспецифическими и не требуют длительного периода для их индукции. Это механизмы врожденного иммунитета, которые действуют немедленно и являются наиболее ранним ответом на инфекцию. Эта ранняя фаза ответа на инфекцию помогает сохранять ее под контролем до тех пор, пока антигенспецифические лимфоциты активируются [89]. Кроме того, продукция цитокинов в течение этой ранней фазы играет важную роль в последующем развитии специфического иммунного ответа (Т-клеточно-опосредованного или гуморального). В случае, если возбудители проходят эти ранние линии защиты, то запускаются механизмы адаптивного иммунного ответа с развитием антигенспецифических эффекторных клеток, специфически распознающих антиген и лимфоцитов памяти, которые длительно сохраняются в организме и предупреждают развитие инфекции, вызываемой повторным попаданием того же самого патогена. Несколько дней требуется для клональной селекции и дифференцировки нативных лимфоцитов в эффекторные Т-клетки и антитело-продуцирующие плазматические клетки. В течение нескольких дней формируется специфическая иммунологическая память, которая обеспечивает длительную защиту при ре-инфицировании тем же возбудителем [89]. В настоящее время классификация типов иммунитета включает в себя: I. Врожденный иммунитет. II. Приобретенный (адаптивный) иммунитет, который, в свою очередь, разделяется на: 1) естественный приобретенный, возникающий после переболевания за счет сохранения клеток памяти. При повторном заражении тем же возбудителем иммунная система быстро реагирует на него именно за счет лимфоцитов памяти, и заболевание может не развиться; 2) искусственный приобретенный, который подразделяется на: а) пассивный – за счет введения в организм готовых антител (например, трансплацентарно от матери к ребенку или с помощью сывороток и препаратов иммуноглобулинов); этот тип защиты быстрый и эффективный, но сохранятся лишь короткое время; б) активный – достигается с помощью безопасных и эффективных вакцин. 2.1. Виды инфекционных агентов и их локализация в организмеВсе известные инфекционные агенты, вызывающие заболевания у человека, разделяются на 5 групп: вирусы, бактерии, грибы, протозойные, гельминты. Их можно обнаружить в разных участках тела. Уничтожение их происходит с помощью разных механизмов иммунологической защиты хозяина. С патогенетической точки зрения важно разделение инфекционных агентов по месту их преимущественного роста и размножения. С этих позиций выделяют 2 основные группы микроорганизмов: 1. Интрацеллюлярные (внутриклеточные). Следует отметить, что фактически все патогены проходят экстрацеллюларную фазу, где они становятся уязвимы для действия антител. В то же время интрацеллюларная фаза развития микроорганизмов остается недоступной для антител и в этом случае основная роль в защите отводится атаке Т-лимфоцитов. 2.2. Виды иммунного ответа на первичную инфекциюПротективный ответ на первичную инфекцию обеспечивается в основном за счет следующих механизмов (Таблица 2) Таблица 2. Преимущественная локализация микроорганизмов и основные механизмы иммунитета [89]
Как видно из этих данных, механизмы протективного иммунитета различны при разных инфекционных заболеваниях, что следует учитывать не только при назначении этиотропной терапии, но и при проведении специфической профилактики, в частности, при вакцинации. Следует подчеркнуть, что в элиминации внеклеточных, в основном пиогенных микроорганизмов (пневмококки, стафилококки, стрептококки и др.) главная роль принадлежит триаде: нейтрофилу, иммуноглобулину и комплементу, причем гибель микроба происходит в нейтрофиле, а комплемент и иммуноглобулины (опсонины) усиливают этот процесс. В элиминации внутриклеточных возбудителей главная роль так же принадлежит триаде: Т-лимфоцитам, NK-клеткам и макрофагам, причем все эти 3 группы клеток обладают способностью синтезировать цитокины (IFN-g, ИЛ-1, ИЛ-2, ФНО-a, и др.), резко усиливающих их функциональные свойства [17]. В ответе на впервые попавшую в организм человека инфекцию выделяют 3 фазы иммунного ответа, отличающиеся как механизмами распознавания антигена, так и эффекторными механизмами. 2.2.1. Фазы иммунного ответаПри первом попадании возбудителя в организм человека включаются механизмы врожденного и адаптивного иммунного ответа (табл. 3). Таблица 3. Фазы иммунного ответа [89]
Как видно из представленной таблицы, адаптивный иммунитет имеет место позже, т.к. антиген-специфические клетки должны подвергнуться клональной селекции, перед тем как они дифференцируются в эффекторные клетки. В соответствии со временем возникновения выделяют следующие фазы:
В случае повторного попадания того же самого возбудителя, т.е. при реинфицировании, удаление возбудителя может происходить с большей скоростью за счет сохранения протективного иммунитета и иммунологической памяти. Протективный иммунитет – это ответ на реинфицирование путем распознавания антигена предсуществующими антителами (преформированными) и эффекторными Т-клетками с последующим удалением возбудителя. Иммунологическая память – проявляется в том, что в ответ на реинфицирование происходит распознавание антигена с помощью В- и Т-лимфоцитов памяти. Это приводит к быстрой экспансии антигенспецифических лимфоцитов, дифференцировке их в эффекторные клетки и удаление инфекционного агента. 2.3. Течение острой инфекции
2.3.1. Механизмы разрушения тканей патогенамиЛюбая "естественная инфекция" сопровождается воздействием на клетки и ткани больного. В результате такого воздействия патогенов на организм происходит разрушение клеток и тканей, которое может осуществляться либо путем прямого воздействия, либо косвенно с привлечением факторов иммунной защиты. Рассмотрим возможные механизмы разрушения [89]: Прямое разрушение тканей патогенамиЭтот процесс происходит за счет: а) продукции экзотоксинов. Экзотоксины, выделяемые микроорганизмами, действуют на поверхность хозяйской клетки, связываясь с соответствующими рецепторами и проникая внутрь клетки, разрушают ее. Примеры: Streptococcus pyogenes, Staphylococcus aureus, Corynebacter diphteriae, Clostridium tetani. Указанные возбудители могут вызвать различные заболевания, такие как тонзиллиты, токсический шок, дифтерию, столбняк; б) выделение эндотоксинов. Эндотоксины, выделяемые при разрушении ряда возбудителей, являются триггерами для синтеза макрофагами цитокинов, которые в свою очередь вызывают локальные или общие симптомы. Примеры: E.coli, Haemophilis influenzae, Shigella, Pseudomonas aeruginosa, Yersinia pestis. Перечисленные возбудители являются этиологическими факторами в развитии сепсиса, менингита, тифа, инфекции органов дыхания, чумы; в) прямого цитопатического действия. Некоторые микроорганизмы способны оказывать прямое разрушающее действие на клетки. В качестве примеров таких возбудителей можно привести: Variola, Varicella zoster, Hepat. B virus, Polio-virus, Measles virus, Influenza virus, Herpes simplex. В результате инфицирования у человека развиваются гепатит В, полиомиелит, корь, подострый склерозирующий панэнцефалит, грипп, герпетическое поражение. Непрямое разрушение тканей патогенамиАдаптивный иммунный ответ к инфекционным агентам может сопровождаться образованием иммунных комплексов, синтезом перекрестно-реагирующих антител, которые связываются с хозяйскими тканями, а также индукцией Т-клеток, разрушающих инфицированную клетку человека. Это все приводит как к удалению возбудителя, так и к разрушению клеток хозяина. Укажем лишь некоторые примеры подобного действия (табл. 4): Таблица 4. Некоторые механизмы повреждения тканей факторами иммунной системы [89]
Как видно из представленных примеров, переболевание так называмыми "естественными" инфекциями, часть из которых относится к вакцин-управляемым, может в ряде случаев либо вызвать, либо поддерживать патологические процессы в организме человека. Таким образом, при рассмотрении вопроса о возможности нежелательных реакций вакцинации следует помнить, что переболевание вакцинуправляемой инфекцией несет в себе не меньшую, а значительно большую опасность в плане разрушающего воздействия на органы и ткани больного. 2.4. Врожденный иммунитет (неадаптивная защита)Первая фаза защиты человека от инфекции, называемая врожденный иммунитет, включает [89]:
Активация комплемента альтернативным путем и захват микроорганизмов тканевыми макрофагами имеют место в ранние часы после инфицирования. Далее включаются механизмы адаптивной защиты – гуморальный и клеточно-опосредованнный иммунный ответ. Ранний неадаптивный ответ важен по 2-м причинам. Во-первых, дает возможность контро-лировать инфекцию до развития адаптивного ответа, он развивается быстро, так как не требует клональной селекции лимфоцитов и, следовательно, не требует латентного периода, как это происходит при пролиферации лимфоцитов и дифференцировке их в эффекторные клетки. Во-вторых, ранний ответ в дальнейшем влияет на адаптивный ответ за счет выработки цитокинов макрофагами. Таким образом, основные отличия врожденного иммунитета от адаптивного следующие: – начинает действовать немедленно после инфицирования; 2.4.1. Основные клетки врожденного иммунитетаК основным клеткам, реализующим механизмы врожденного иммунитета, относятся макрофаги, нейтрофилы, естественные киллеры. Макрофаги происходят из циркулирующих в крови моноцитов. В большом количестве они находятся в соединительной ткани, селезенке, печени. Это большие фагоцитирующие клетки, которые играют ключевую роль во всех фазах защиты человека от инфекции. В отличие от моноцитов на макрофагах имеются рецепторы к маннозе, а также СД14-рецептор, который предназначен для связывания с бактериальными липополисахаридами. После связывания бактерий с рецепторами макрофагов происходит инициирование синтеза цитокинов макрофагами, а поглощенные бактерии перевариваются. В свою очередь, медиаторы воспаления рекрутируют новые фагоцитарные клетки в места локализации инфекции. В частности, макрофаги, активированные с помощью ЛПС (липополисахариды) бактерий, после распознавания, захвата и дегрануляции Гр(–) бактерий синтезируют интерлейкины – ИЛ-1, ИЛ-6, ИЛ-8, ИЛ-12 и TNF-a. Другие медиаторы, выделяемые макрофагами в ответ на инфекционные агенты, – это простагландины, оксид азота (NO), лейкотриены (в частности, LB4), PAF-фактор, активирующий пластины. В свою очередь, эти продукты активируют комплемент – С5а, С3а и в меньшей степени С4а. Один из компонентов комплемента, а именно С5а, – активирует тучные клетки, что приводит к выделению гистамина, серотонина и влиянию на эндотелиальные клетки в зоне воспаления. Все указанные факторы участвуют в воспалении. Рассмотрим более подробно локальные и системные эффекты монокинов – цитокинов, выделяемых макрофагами (рис. 1). 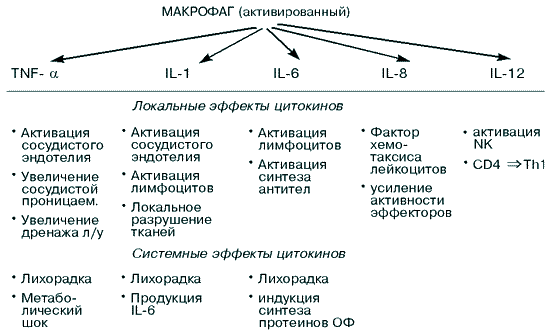 Рис. 1. Эффекты монокинов [89] Как уже отмечалось, макрофаги поглощают и разрушают в основном Гр(–) бактерии, при этом активируются с помощью ЛПС бактерий и вследствие этого начинают секретировать монокины – интерлейкины 1, 6, 8, 12 и TNF-a. Монокины, выделяемые макрофагами в ответ на бактериальную инфекцию, вызывают следующие эффекты: – индуцируют продукцию протеинов острой фазы печенью; Помимо макрофагов, в реализации механизмов врожденного иммунитета большую роль играют нейтрофилы. Эти клетки преобладают в раннем клеточном инфильтрате в месте воспаления. Они находятся в крови и практически отсутствуют в нормальных тканях. Нейтрофилы – это коротко живущие клетки. В среднем срок их жизни равен 3-4-м дням. При реализации врожденного иммунного ответа продуцируются разные факторы, часть из которых является хемоаттрактантами для нейтрофилов и они быстро эмигрируют из крови и входят в очаг инфекции. В очаге нейтрофилы способны элиминировать многие патогены путем фагоцитоза. Нейтрофилы способны фагоцитировать как опсонизированные бактерии, т.е. в соединении с антителами (опсонинами), так и в отсутствии антител. Таким образом, нейтрофилы осуществляют первую линию защиты. Компоненты бактериальной клетки могут быть связаны напрямую с нейтрофилами или опосредовано через липополисахариды – LPS, которые вначале связываются с протеинами сыворотки, образуя липополисахаридсвязанный протеин – LBP. Комплексы LPS и LBP затем соединяются с СД 14 на поверхности нейтрофила. Нейтрофилы могут также фагоцитировать комплексы бактерий, соединенные с С3b компонентом комплемента. Это соединение в свою очередь инициирует альтернативный путь активации комплемента. Нейтрофилы способны элиминировать Гр(–) и Гр(+) бактерии, грибы и даже некоторые оболочечные вирусы. В то же время многие нейтрофилы погибают в очагах воспаления. Часть микроорганизмов фагоцитируется нейтрофилами, но не убивается ими. Но даже такая секвестрация микроорганизмов важна для хозяйской защиты, так как позволяет контролировать инфекцию в ранний период, а секвестрированные частицы вновь рефагоцитируются другими нейтрофилами или макрофагами, которые аккумулируются в зоне воспаления. Из сказанного следует, что нейтрофилы – это важная составная часть врожденного иммунного ответа хозяина на инфекцию. Естественные киллеры (NK-клетки) служат в качестве самой ранней защиты против интрацеллюлярных микроорганизмов. NK-клетки были идентифицированы по их способности убивать определенные опухолевые клетки in vitro без предварительной активации. Однако известна их роль в ранней фазе защиты от инфицирования интрацеллюлярными патогенами, особенно Herpes virus и Listeria monocytogenes. Активность естественных киллеров увеличивается в 20-100 раз, когда на них воздействуют цитокины, в частности, IFN-a, IFN-b или IL-12. Последний называется также NK-активирующим монокином, который продуцируется в раннюю фазу инфекции. ИЛ-12 в сочетании с TNF-a могут вызвать усиление продукции IFN-g естественными киллерами. Поскольку NK-клетки опосредуют в основном защиту человека против вирусов, они должны иметь некие механизмы для дифференцировки инфицированной и неинфицированной клетки. Окончательно этот вопрос не решен, однако установлено, что NK-клетки избирательно убивают клетки-мишени, имеющие низкий уровень молекул MHC класса 1 на своей поверхности. Это предполагает следующий механизм действия NK-клеток [89]: молекулы МНС класса 1 на нормальных клетках распознаются NK-клетками с помощью рецептора Ly49. В результате такого распознавания посылается отрицательный сигнал и происходит ингибиция активации NK-клетки. Иными словами, если рецептор Ly49 на естественных киллерах распознает молекулы МНС класса 1 на поверхности хозяйской клетки, то происходит ингибиция активации NK-клеток и последующего киллинга клетки-мишени не происходит. Если клетки-мишени не экспрессируют молекулы МНС класса 1, то происходит киллинг их и клетка-мишень погибает. IFN-a и IFN-b усиливают экспрессию молекул МНС класса 1 на поверхности нормальных неинфицированных клеток, тем самым предотвращая атаку на них NK-клеток. Вирус-инфицированные клетки экспрессируют значительно меньшее количество молекул МНС класса 1, либо на их поверхности отсутствуют эти молекулы. В ряде случаев молекулы МНС класса 1 становятся измененными. В этих случаях рецептор Ly49 не распознает молекулы МНС класса 1 (НLA-B аллели) и нет подачи негативного сигнала. В результате NK-клетка активируется и убивает клетку-мишень. Активация NK-клетки происходит через поверхностный рецептор NK R-P1 – это лектин-подобный рецептор, распознающий карбогидратные молекулы, являющийся триггером киллинга NK-клеток. Таким образом, вирусинфицированные клетки могут быть подвергнуты киллингу с помощью NK-клеток двумя путями: – если вирусы ингибируют весь синтез протеинов хозяина, и тогда а NK-клетки убивают клетку-мишень, т.к. нет подачи негативного сигнала через рецептор Ly49; С другой стороны, известно, что вирус-инфицированная клетка распознается NK-клеткой даже, когда МНС экспрессия не нарушена вирусом. Люди с дефицитом NK-клеток очень чувствительны к ранней фазе герпес-инфекции [89]. Подводя итоги, следует сказать, что ранний неадаптивный ответ (врожденный иммунитет) на инфекцию вовлекает широкий круг эффекторных механизмов, направленных на разные классы патогенов. Все ответы становятся возможными благодаря рецепторам, которые являются в основном неспецифичными.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||